Дальнейшие доказательства устойчивости систем переноса углеводов в клетки психрофилов при температурах (0—10°) были получены Чирилло с сотрудниками (Cirillo et al., 1963) при сравнении температурных характеристик поступления глюкоз- амина и сорбозы в клетки психрофильного штамма Candida sp. и мезофильного Saccharomyces cerevisiae. Значения температурной характеристики (μ) составляли 9 и 52 ккал/моль для аккумуляции глюкозамииа и 12 и 49 ккал/моль для сорбозы у психрофила и мезофила, соответственно. На рис. 36, где представлены данные, касающиеся транспорта L-сорбозы, метаболически инертного и поэтому очень удобного «модельного сахара», отличие психрофилов по реакциям их транспортных систем на низкие температуры очень хорошо заметно. Температурные ко- эффициенты процесса Q10 (1—10°) равны 5,0 для ме- зофила и 1,74 — для психрофила. Корреляция температурных характеристик роста и транспорта углеводов позволяет сделать заключение, что минимальная температура роста организма определяется вероятнее всего температурной характеристикой системы транспорта углеводов.
К таким же выводам привели и эксперименты по определению температурной зависимости процесса накопления Д-глюкозамина у психрофильных штаммов Arthrobacter и Candida, которые хорошо росли при 0° и активно накапливали глюкозамнн в клетках, тогда как у родственных мезо- фильных штаммов эти процессы прерывались при 5 и 10°. Однако эта разница в чувствительности транспорта у психрофилов и мезофилов не сказывалась на реакции их систем на ингибиторы. Были проверены разобщающий яд динитрофенол (2,4-ДНФ), снижающий энергетический потенциал клетки и отключающий энергию от переноса углеводов (Гершанович, 1973), уранилацетат, ноны UO+2 которого соединяются, как полагают, с фосфатными группами в фосфолипидных молекулах цитоплазматической мембраны, и нистатин, ингибирующее действие которого основано на соединении со стеролами той же мембраны. Эффективность 2,4-ДНФ свидетельствовала о том, что поступление углеводов в клетки обоих типов организмов является активным транспортом, требующим затрат энергии, и, следовательно, о сопряжении реакции переноса с энергетическим обменом. Блокирование фосфатных групп в мембране, создающее трудности в функционировании пермеаз, таклее инактивирует субетраттранспортные системы изучаемых организмов. Сами авторы, Роуз и Эвисон (Rose, Evison, 1965), отдают предпочтение гипотезе низкотемпературной инактивации у мезофилов этих белков-переносчиков.
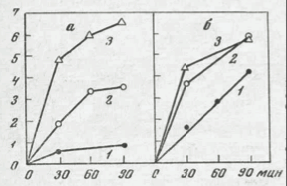
Рис. 36. Зависимость транспорта сорбозы в клетки Saccharomyces cerevisiae (а) и Candida (б) от температуры (Cirillo et al„ 1963) 1—1°, 2— 10°, 3—30°, на оси ординат — концентрация сорбозы, мг/мл клеточной воды