Нарушения в синтезе белка могут быть первичным фактором нарушения общего метаболизма клеток, приводящего к инактивации и отмиранию организма, или следствием термоиндуцированного дефекта в синтезе РНК.
Первая возможность реализована, например, у облигатного психрофила Vibrio marinus (Cooper, Morita, 1972). Кроме того, она хорошо представлена и подробнее, чем у других объектов, изучена у Micrococcus cryophilus (Malcolm, 1968а, b). Было замечено, что угнетение включения С14-урацила (т. е. синтеза РНК) начинается после термоиндуцированного (30°) прекращения синтеза белка, и между этими двумя моментами лежит заметный промежуток времени. Одновременно наблюдалось значительное возрастание выхода из клеток нуклеотидов. Полагают, что это были продукты деградации уже существующей РНК. Способ, посредством которого повреждение в синтезе белка сказывается на синтезе РНК у этого организма, неизвестен. Высказываются соображения о возможном функционировании механизма обязательного аминокислотного контроля над синтезом РНК, допускаемого для ряда объектов (Neidhardt., цит. по Malcolm, 1968b). В этом случае накопление РНК должно зависеть от связывающей активности аминоацил-тРНК-синтетаз, хотя истинный биохимический механизм остановки накопления тРНК в присутствии поврежденной комплементарной синтетазы неясен, как и принцип контроля в целом).
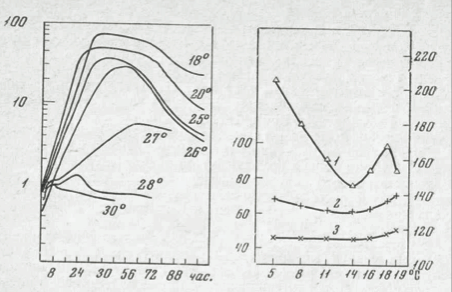
Рис. 29. Действие температуры инкубации на содержания РНК в культурах Micrococcus cryophilus (Gray, Jackson, 1973)
На оси ординат — содержание рибозы, мкг/мг
Рис. 30. Действие температуры на содержание РНК (1), белка (2) и ДНК (3) в культурах психрофильного штамма Pseudomonas sp. (Harder, Veldkamp,1967)
На оси ординат: слева — содержание белка, мг/мг сухого вещества клеток, Х100 и ДНК мкг/мг сухого вещества клеток, справа — содержание РНК, мкг/мг сухого вещества клеток