Продолжение этих работ [Жданова и др., 1978а] показало, что УФ-облучение конидий грибов и изолированного пигмента при низких температурах (<00) вызывает увеличение сигналов ЭПР.
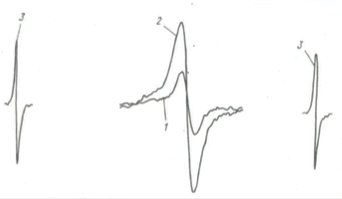
Рис. 8. Фотоиндукция сигнала ЭПР в растворе аспергиллина (конидиального пигмента Aspergillus niger) [Rezska, Pajak, 1975]
1 — темновой сигнал ЭПР; 2 — сигнал ЭПР после облучения; 3 — сигнал ЭПР внутреннего стандарта.
Если облучение проводилось в течение достаточно длительного отрезка времени, то быстрый вначале рост сигнала ЭПР замедлялся и приближался к некоторому предельному значению, зависящему от вида гриба, содержания меланина в его клеточных стенках, состава атмосферы, в которой проводилось облучение, длины волны света (λ=250-390 нм, светофильтр УФС-1 и λ≤700 нм, светофильтр КС—18) и температуры (О, -40, -80 и -120°). Меланины УФ-чувствительных штаммов мутанта К-1 Cl. transchelii и Oidiodendron cerealis, для которых характерно пониженное содержание ПМЦ, больше изменялись под действием облучения, чем пигменты УФ-резистентных грибов. После прекращения облучения фотоиндудированный при -120° сигнал в течение 1 часа уменьшался незначительно (за исключением сигнала от мутанта К-1, который за первые 15 мин терял 25% интенсивности). При -80 и -40° эффект ослабления фотоиндуцированного сигнала был выражен сильнее (в возрастающей степени), а при 0° скорость гибели фотоиндуцированных ПМЦ была столь велика, что даже при постоянном облучении сигнал ЭПР не отличался от исходного. С меланинами, выделенными из клеток (фракция 3б из Cl. transchelii), циклы заморажение (-100- 120°) — облучение видимым светом — разморажение до комнатной температуры (с возвратом сигнала ЭПР к исходному уровню) можно было повторять многократно: все эффекты были полностью обратимы.
Авторы считают, что ПМЦ в меланинах генерируются по одиоквантовому механизму, активен свет с λ<900 нм. При этом в пигменте происходят процессы переноса электронов, приводящие к образованию анион- и катионрадккальных центров — «электронов» и «дырок». Полупроводниковые свойства меланина обеспечивают обратимость этих процессов и предотвращают необратимые химические изменения пигмента, защищающего клетку от действия УФ- и видимого света. Отсутствие регистрируемого повышения сигнала ЭПР во время облучения пигмента при температурах ≥0° авторы связывают с очень быстрой темновой рекомбинацией индуцированных ПМЦ, отрицая возможность «выключения» при повышении температуры первичных фотофизических процессов, генерирующих ПМЦ. Они считают, что и в природе в условиях обычных, «экологические» температур свет (λ≤800 нм), действуя на меланинсодержащие структуры грибов, вызывает в них процессы фотопереноса электронов, сопровождающиеся темновой рекомбинацией образовавшихся анион- и катион-радкальных центров.