Тенденция к усилению синтеза ненасыщенных жирных кислот как генотипическое или модификационное приспособление к низким температурам достоверно показано на многих, но не на всех исследованных организмах (Гельман и др., 1972; Александров, 1975; Schmidt-Lorenz, 1967b, 1972; Kaneda, 1971).
В связи с тем, что этот вопрос подлежит специальному рассмотрению в главе V, здесь мы только попытаемся представить себе крайне упрощенно, конечно, какие последствия для низкотемпературного функционирования мембран может иметь это изменение в жирнокислотном составе липидов.
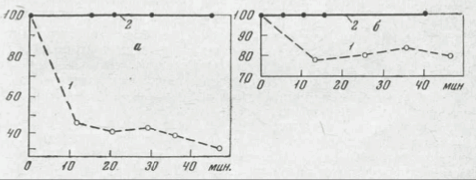
Рис.17 Действие продолжительности холодового шока на жизнеспособность мезофильного (а) и психрофильного (б) штаммов. Pseudomonas в зависимости от температуры предннкубации (Farrel, Rose, 1968а).
1-охлаждение от 30° до -2°, 2-ох- лаждение от 10° до -2°. на оси ординат — выживаемость, %
При этом следует иметь в виду, что большинство имеющихся данных характеризуют общий липидный состав микробных клеток. Только у грамположительных бактерии большая часть липидов клетки (до80%) локализована в цитоплазматической мембране и других мембранных образованиях. У грамотрицательных бактерий картина осложняется присутствием этих веществ в клеточной стенке (Гельман и др., 1972), что, впрочем, при обсуждении интересующего нас вопроса не должно создавать особых помех из-за анатомической и функциональной связи этих структур. Хуже обстоит дело с привлечением данных по общим липидам дрожжей и грибов, у которых запасные формы могут значительно преобладать над структурными.